11 Физиологическая организация мигрени
11
Физиологическая организация мигрени
Двадцать четыре буквы не способны составить большее разнообразие слов на всевозможных языках, чем разнообразие симптомов, производимое меланхолией у нескольких пациентов. Эти симптомы нерегулярны, темны, разнообразны, неисчерпаемы – сам Протей не так изменчив…
Бертон
Проблема мигрени, как и меланхолии, о которой писал Бертон, заключается в бесконечном разнообразии симптомов, каковые мы призваны объяснить. Мы должны попытаться сформулировать теорию или набор теорий, более общих, нежели простые механизмы, разобранные нами в предыдущей главе: теория должна быть общей настолько, чтобы объяснить каждый аспект каждого типа мигрени и чтобы ее можно было тем не менее приложить к каждому частному симптому.
Мы начнем с рассмотрения симптомов мигрени на разных функциональных уровнях – от низшего к высшим, а также с рассмотрения возможных механизмов, отвечающих за эти симптомы. Если такой метод окажется несостоятельным, мы будем вынуждены прибегнуть к идеям, радикально отличающимся от «функции» или «центра» в том смысле, как их понимают в нейрофизиологии. Нам придется мыслить в понятиях динамической организации функциональных систем, а не в традиционных понятиях фиксированного нервного аппарата и механизмов его работы.
Мы уже видели, что вегетативные симптомы мигрени проявляются повышенной активностью парасимпатической нервной системы, и нас не может не удивить тот факт, что выступающие на первый план разнообразные симптомы соответствуют анатомической и функциональной организации парасимпатической нервной системы. Существуют анатомически дискретные ганглионарные сплетения во всех основных висцеральных, сосудистых и железистых структурах организма; они составляют самый нижний уровень нервного представительства внутренних органов тела. Мы можем постулировать, что существование таких функционально дискретных сплетений может обеспечить физиологическую основу разнообразных, отличающихся друг от друга парасимпатических синдромов: локальная нейронная активность в таких сплетениях, запущенная начальным разрядом из центральной нервной системы, может устойчиво поддерживаться – в течение минут, часов или дней – как функционально дискретное, изолированное возбуждение. В случае когда у больного имеет место длительная, локализованная в одной половине головы боль, мы можем наблюдать относительно изолированное расстройство, заключенное в одном околососудистом сплетении; в других случаях местное нейронное возбуждение может ограничиваться интрамуральными сплетениями желудка, толстой кишки, слезных желез и т. д. Первоначальное нейронное нарушение может затем усугубиться вследствие местных тканевых изменений, например транссудации или асептического воспаления, что и происходит при затянувшейся сосудистой головной боли или других долговременных расстройствах, следующих за исходным нейрогенным нарушением. Вегетативные симптомы мигрени на своем последнем и самом нижнем уровне могут передаваться в разных направлениях, менять интенсивность и изолироваться за счет сетевой конфигурации парасимпатической нервной системы. «Инструкции» относительно выбора формы приступа или эквивалента мигрени определяются, вероятно, центральными механизмами, но вспомогательную роль могут играть местные различия в порогах – либо врожденные, либо закрепленные условными рефлексами. Характерно, однако, что, если из системы удалить какой-либо орган-мишень или местное сплетение (например, при хирургическом вмешательстве), мигренозные приступы возобновятся в несколько иной форме. Это позволяет предположить, что периферические механизмы используются в меру их доступности и центральная организация мигрени отличается пластичностью и гибкостью.
Изначально термин «парасимпатический» использовали и продолжают в ограниченном варианте использовать теперь для обозначения периферических структур и периферической активности. Ясно поэтому, что нельзя говорить о мигрени как о парасимпатическом приступе, так как приступ имеет также и центральные, церебральные составляющие. Подходящие концепции и термины были выработаны Гессом на основе его известных исследований центральных вегетативных и диэнцефальных функций (Гесс, 1954). Гесс использует термин «эрготропный» для обозначения связи периферической симпатической активности с центральным возбуждением и термин «трофотропный» для обозначения противоположного явления. Эти термины являются не только физиологическими, но также биологическими и организменными. Так, эрготропия обозначает тенденцию организма нацелить себя на внешний мир, на то, чтобы быть активным, выполнять работу и т. д., для чего нужен повышенный уровень бодрствования и усиление остроты сенсорного восприятия, повышение мышечного и симпатического тонуса и т. д. Трофотропия обозначает тенденцию организма нацелить себя вовнутрь, на сохранение внутреннего баланса, для чего повышается активность внутренних органов и желез, несколько снижается уровень бодрствования, острота сенсорного восприятия и мышечный тонус. Рекомендованные Гессом термины весьма полезны для нашего понимания природы мигренозного приступа. Очевидно, что все его критерии трофотропии (повышение парасимпатического тонуса, снижение возбуждения, синхронизация ЭЭГ и т. д.) выполняются во время мигренозного приступа: основная часть мигрени – это, можно сказать, полиморфный трофотропный синдром.
Экспериментальной разработкой концепций Гесса и их клиническими приложениями занимались многие авторы, среди которых следует особо выделить Гельхорна. Так, им было показано (см. Гельхорн, 1967), что на всех уровнях нервной оси существуют эрготропные и трофотропные системы, различающиеся между собой анатомически, физиологически и фармакологически. Было также показано, что эрготропная и трофотропная активность находятся в антагонистическом взаимодействии, чем поддерживается их равновесие. Это равновесие определяет то, что Гельхорн называл «настройкой» нервной системы в каждый данный момент времени. Так, торможение активности эрготропной системы приводит к возбуждению трофотропного отдела, и наоборот. В типичных случаях, однако, резкие сдвиги вегетативной настройки сопровождаются противоположно направленным рикошетом.
Теперь мы можем перевести последовательность событий при простой мигрени на язык терминов Гесса. Продромальные симптомы, или симптомы-предвестники, проявляются преобладанием эрготропной активности; собственно приступ характеризуется переходом к трофотропии. Симптомы рикошета являются эрготропными. Таким образом, простую мигрень можно представить себе как трехстадийный медленный пароксизм, в ходе которого происходит последовательная и характерная смена настроек. Такая идея приблизительно соответствует представлению Леннокса о том, что мигрень – это «вегетативный припадок». Мы не можем точно установить уровень, на котором начинается мигренозный процесс, и можно думать, что такая постановка вопроса вообще лишена смысла, ибо эрготропная и трофотропная системы представлены иерархически на всех уровнях нервной оси – от боковых рогов спинного мозга до ретикулярной формации ствола головного мозга, гипоталамуса и, наконец, медио-базальных отделов мозговой коры.
Цикл возбуждения и торможения можно выявить при самом компактном течении мигренозной ауры: цикл возбуждения (мерцающая скотома, парестезии, возбуждение, диффузное усиление остроты сенсорного восприятия и т. д.) сменяется циклом торможения (негативная скотома, анестезия, заторможенность, обмороки, синкопы, снижение остроты сенсорного восприятия и т. д.), а затем (не всегда) следует рикошетное повторное возбуждение длительностью не более 30–40 минут.
Таким образом, мы возвращаемся к картине мигренозного процесса, предложенной Лайвингом столетие назад: форма центрального мозгового припадка, активность которого проецируется рострально в направлении полушарий, а периферически через разветвления вегетативной нервной системы. Можно представить себе кору головного мозга как объект восходящей импульсации в течение мигренозной ауры; на эту импульсацию кора отвечает своим вторичным возбуждением: эта вторичная активность является многофокусной (мерцающая скотома, парестезии и т. д.) и протекает на фоне диффузного возбуждения коры. Аналогично мы можем рассматривать периферические вегетативные сплетения как объекты нисходящей импульсации, на которую сплетения отвечают вторичной многофокусной активностью. Такая картина мигренозного процесса схематически представлена на рисунке 7.
Очевидно, однако, что нам потребуются другие концепции и термины, чтобы описать активацию мозговой коры во время мигренозной ауры. Зрительные галлюцинации при мигрени дают нам отчетливые указания на эти высшие процессы и на их организацию.
Мы уже видели (см. главу 3), что существует тенденция к последовательности зрительных галлюцинаций – от наиболее простых и элементарных до невероятно сложных. Эта последовательность очень напоминает ответ на определенные биологически активные вещества (например, мескалин), лишение сна или сенсорную депривацию. Так, мы можем сравнить последовательность событий при мигрени с сообщением Хебба (1954) о зрительных галлюцинациях, вызванных сенсорной депривацией:
«Представляется, что активность совершает в своем развитии весьма упорядоченный переход от простого к сложному. Первый симптом: при закрытых глазах поле зрения меняет цвет, становясь из темного светлым. Затем испытуемый сообщает о появлении точек, линий и простых геометрических фигур… На следующем этапе он уже видит что-то похожее на рисунок обоев. Потом появляются отдельные предметы, лишенные фона… и, наконец, испытуемый видит связные сцены с характерными для сновидений искажениями».
Формы и перемещения простейших мигренозных фосфенов в поле зрения напоминают цветовые и абстрактные галлюцинации (мелькающий свет, звезды, колеса, диски, крутящиеся перила и т. д.), описанные Пенфилдом и Расмуссеном при прямом раздражении поверхности коры головного мозга (поле 17).
Феномен мерцающей скотомы уникален именно для мигрени, и ее пока не удалось воспроизвести ни в одном эксперименте. Лэшли (1941) предположил, что характеристическая микроструктура этих скотом (край, зазубренный мелкими углами, более грубая текстура в нижней части поля зрения, как показано на рисунке 3) соотносится с цитоархитектоникой зернистого слоя первичной зрительной коры.
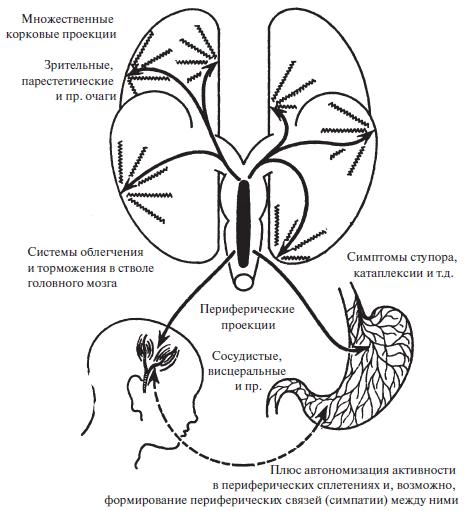
Рис. 7. Схема гипотетического мигренозного процесса. Схематическое представление мигренозного процесса как медленной, циклической центрально-мозговой пароксизмальной активности, которая рострально проецируется к коре головного мозга, где возбуждает вторичные процессы мигренозной ауры (скотомы, парестезии и т. д.), и каудально, к разветвлениям вегетативных сплетений по всему организму. Этот процесс (выражаясь словами Говерса) является «…очень загадочным… это особая форма активности, которая распространяется, как круги по поверхности пруда, в который бросили камень… в участке, по которому прошла волна, остается нечто вроде молекулярного возмущения».
Лэшли тоже высказался по поводу скорости перемещения скотом и вычислил, что она соответствует волне возбуждения, перемещающейся в первичной зрительной коре со скоростью около 3 мм в минуту. За волной возбуждения следует волна торможения. Лэшли признает, что «ничего не известно о процессах, лежащих в основе этой нервной активности», и потому не предлагает теории, объясняющей эту активность либо местным корковым нарушением, либо возбуждением, пришедшим из подкорковых структур.
Неясно, почему зрительная кора проявляет большую чувствительность к стимуляции, чем соответствующие области кожно-кинестетической коры (поле 3) или слуховая кора (поле 41); непонятно также, какие фундаментальные процессы определяют частоту мерцания скотом или парестезий (6—12 в секунду). Но, вероятно, это не простое совпадение, что частота мерцаний соответствует частоте альфа-ритма и частоте стробоскопического мелькания, вызывающей стимуляцию на ЭЭГ, припадки фотоэпилепсии и световые скотомы. Можно предполагать, что данная частота является фиксированной мозговой частотой, предназначенной для переработки информации и сканирования объектов.
Вслед за появлением простых фосфенов и мерцающих скотом могут последовать зрительные нарушения и галлюцинации более высокого порядка: микропсия и макропсия, различные формы зрительной агнозии, мозаичное зрение и стереотипированные (кинематические) последовательности зрительных образов. Экспериментально доказано (Пенфилд и Расмуссен, 1950), что непосредственная стимуляция вторичных и периферических полей зрительной коры может вызывать появление сложно организованных галлюцинаций, выстроенных в определенной пространственно-временной последовательности, и можно предположить, что все более сложные зрительные галлюцинации возникают на этом же уровне (или выше). Что же касается мозаичного зрения и различия в размерах мозаичных фрагментов (см. рис. 4), то здесь можно думать о какой-то форме функциональной схематизации, которая происходит за пределами анатомически фиксированных цитоархитектонических единиц.
Конорский (1967) выдвинул и обосновал теорию перцептивных или гностических единиц (минимальных перцептивных структур), которые могут играть роль в появлении мозаичности зрения. Мы уже указывали, что вначале мозаичность проявляется зернистостью изображения, впечатлением, что изображение состоит из мелких кристаллов, затем мозаика становится грубее, и так продолжается до наступления полной агнозии – то есть изображения становятся неузнаваемыми. Можно предположить, что эти симптомы представляют собой субъективное отражение прогрессивного увеличения гностических единиц, в норме невидимых, но начинают осознаваться по мере увеличения, представляясь в виде полигональных структур, становящихся с увеличением размеров все более грубыми. Процесс продолжается до тех пор, пока размер не начинает превышать информативное изображение, и в этот момент распознавание предмета становится затрудненным или невозможным, как распознавание предмета на фотографии со слишком крупным зерном.
Наиболее сложные сенсорные галлюцинации мигренозной ауры принимают форму синестезии и других сенсорных взаимодействий, а также форму сноподобных последовательностей с появлением сенсорных образов любой модальности, сенсорной и моторной афазии, а также общих расстройств мышления и поведения.
Можно далее предположить, что иерархия галлюцинаций мигренозной ауры коррелирует с последовательной активацией различных корковых полей. Центральные корковые поля (например, поле 17 зрительной коры) «отличаются от остальных полей “грубостью” нейронной структуры, приспособленной для восприятия и генерации мощных волн возбуждения» (Лурия, 1966). Эти центральные поля, а в особенности поля зрительной коры в наибольшей степени чувствительны к распространяющемуся в ростральном направлении возбуждению, а ответ проявляется простыми соматотопическими галлюцинациями в зрительных или тактильных полях (в виде скотом или парестезий). Более сильное возбуждение может достигать вторичных сенсорных полей, и здесь возникает уже агнозия и более сложные галлюцинации какой-либо определенной модальности. При наиболее сильной стимуляции возбуждаются третичные сенсорные поля, и тогда (пользуясь словами Лурия) возникают «наиболее сложные формы интеграции объединенной активности зрительного, слухового и кинестетического анализатора». Эта стимуляция приводит к дезорганизации корковой активности, то есть к сложным нарушениям состояния сознания. На рисунке 8 обозначены области коры, соответствующие первичным, вторичным и третичным полям, их перекрывания и границы между ними.
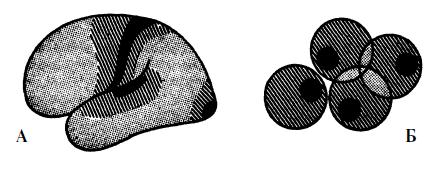
Рис. 8. Корковые поля, имеющие отношение к возникновению мигренозной ауры. Распределение и границы первичных (черный цвет), вторичных (затененные области) и третичных (слегка заштрихованные участки) полей коры головного мозга. Элементарные сенсорные галлюцинации (зрительные, тактильные, слуховые) возникают, предположительно, вследствие возбуждения низкопороговых первичных полей, а более сложные нарушения перцептивных и интегративных функций возникают в результате распространения интенсивного возбуждения на вторичные и третичные поля (рисунки выполнены Поляковым).
Мы видим, что при мигрени очень широк диапазон нарушений, простирающийся от элементарных вегетативных нарушений (с вовлечением периферических вегетативных сплетений) и нарушений центральных механизмов возбуждения до разнообразных корковых расстройств с вовлечением корковых полей нескольких нейронных уровней. Таков репертуар мигрени. Теперь мы займемся вопросом о многообразии симптомов и проявлений внутри этого диапазона. При простой мигрени кора головного мозга не участвует в формировании клинической картины. При изолированной мигренозной ауре проявляются главным образом именно корковые симптомы. При классической мигрени проявления касаются нескольких уровней.
Нам придется вникнуть в физиологические основы превращений и трансформаций, столь характерных для мигрени: переходов от одного мигренозного эквивалента к другому, от простой мигрени к классической, от мигрени к эпилепсии, обморокам, вагальным приступам и всем прочим реакциям, которые Говерс называл пограничными. Именно эти реакции являются самыми трудными и интересными аспектами мигрени, и совершенно ясно, что к ним невозможно подойти, если не взять на вооружение радикально иную концепцию функции головного мозга. Невозможно объяснить меняющиеся уровни и трансформации мигрени в понятиях фиксированных нейронных механизмов.
Это отчетливо понимали уже сто лет назад Лайвинг и Хьюлингс Джексон. Оба дали похожие ответы, хотя ответ Джексона представляется более отработанным и ясным. Лайвинг писал, что «…концентрация этой тенденции (нервной силы) в определенных местах… главным образом и определяет характер обсуждаемого невроза». Джексон был вынужден отказаться от представлений о единых анатомических центрах и анатомически локализованных функциях и вместо этого представить себе нервную систему, как систему организованную иерархически, состоящую из нескольких уровней, на каждом из которых представлена каждая функция:
«Я полагаю, что нервная система – снизу доверху – представляет собой сенсорно-двигательный механизм; каждая часть нервной системы представляет либо восприятие, либо движение, либо и то и другое. Периферия – это самый нижний уровень; но мы будем говорить о трех уровнях центральной эволюции. (1) Самый нижний уровень состоит из передних и задних рогов спинного мозга и их гомологов в более высоких отделах мозга. (2) Средний уровень состоит из двигательной области Ферье с ганглиями полосатого тела, а также из чувствительной области. Таким образом, организм независимо дважды представлен в нервной системе. (3) Самый высший уровень состоит из высших двигательных центров (префронтальная область) и высших сенсорных центров (затылочные доли)».
Таким образом, каждая функция должна иметь сложную «вертикальную» организацию: локализацию симптома ни в коем случае нельзя идентифицировать с локализацией поврежденной функции. При различных типах эпилепсии наблюдается распад функции: так большой припадок рассматривается как патология высшего уровня, а приступ ларингоспазма (ларингеальная эпилепсия или мигрень) считается патологией низшего уровня. Джексон касается мигрени лишь вкратце, но понятно, что он имеет в виду только классическую мигрень, когда пишет:
«Я уверен, что эти случаи мигрени являются случаями эпилепсии [сенсорной эпилепсии]. Думаю, что сенсорные симптомы пароксизма возникают благодаря “разрядному повреждению” путей, ведущих от зрительного бугра, то есть от “сенсорного центра среднего уровня”. Я уверен, что головная боль и рвота возникают уже после пароксизма».
Простая мигрень с ее преимущественно вегетативной симптоматикой должна быть отнесена к припадкам низшего уровня. Мигренозные ауры со сложными галлюцинациями и состояниями помрачения сознания должны быть отнесены к припадкам высшего уровня. Все формы мигрени, по мнению Джексона, отличаются одинаковой организацией, осуществляемой идентичными механизмами на разных уровнях.
В своем анализе эпилепсии Джексон обращает особое внимание на иерархическое представительство движений и двигательных функций в нервной системе, в то время как нас больше интересует вертикальная организация вегетативных и сенсорных функций. Факт, что и они дублируют свое представительство на все более высоких уровнях нервной оси, предоставляет нам новую координату выбора в оценке протекания приступа мигрени. Мигренозный синдром может проявляться на высшем джексоновском уровне (в виде сложной ауры), на среднем джексоновском уровне (в виде элементарной ауры, вовлекающей только первичные сенсорные поля коры головного мозга) или на низшем джексоновском уровне (в виде простой мигрени или мигренозных эквивалентов). Джексон также говорит о коллатеральном и вертикальном распространении эпилептических симптомов. Коллатеральное распространение – это вовлечение в эпилептический процесс прилегающих участков того же уровня. Отсутствие анатомически определенного «разряжающегося очага» означает, что нет пределов такому коллатеральному распространению. Так, аура, возникающая из-за активации первичных сенсорных полей, может вовлечь или «выбрать» без разбора любое зрительное или тактильное поле, в то время как простая мигрень низшего уровня может вовлечь в процесс любое вегетативное поле на уровне ствола головного мозга, порождая то приступ головной боли, то приступ болей в животе, то приступ боли в груди.
Верификация и расширение идей Джексона относительно функций и их локализации в нервной системе стали предметом особого внимания рефлексологов послепавловской эпохи. В свете новых данных потребовался радикальный пересмотр и переосмысление основных доктрин (см. Лурия, 1966). Так, согласно исследованиям послепавловской школы, на самом деле выполнением определенных биологических задач занимаются соответствующие функциональные системы. Самым значимым отличительным признаком такой функциональной системы является то, что она основана на динамической «констелляции» связей, находится на разных уровнях нервной системы, при этом элементы системы могут замещать друг друга, менять задачи, сами оставаясь неизменными. Таким образом (если воспользоваться словами Лурия), «такая система функционально объединенных компонентов имеет системную, а не конкретную структуру, в которой исходная и конечная связи системы (задача и эффект) остаются постоянными и неизменными, в то время как промежуточные связи (средства представления задачи) могут в широких пределах изменяться.
Эти соображения, обсуждаемые Лурия в отношении к движениям и двигательным задачам, годятся и для понимания мигрени и ее превращений как вегетативной и психосоматической задачи. Задача может иметь вид необходимости нейронного разряда (как в случае периодической или пароксизмальной мигрени) или физической или эмоциональной потребности, при обусловленных или ситуационных типах мигрени. Конечная связь, эффект мигрени – это восстановление физиологического (или эмоционального) равновесия. Но адаптивная (приспособительная) задача имеет системную, а не конкретную структуру, то есть использованные для ее решения механизмы могут быть многочисленны, разнообразны и непостоянны. Видов мигрени и способов ее «исполнения» может быть так же много, как способов приготовления омлета. Если откажет одна промежуточная связь, один промежуточный механизм, то система реорганизуется таким образом, чтобы восстановить способность к решению задачи.
Так же как и в случае двигательных или перцептивных задач, частные механизмы подчинены общей стратегии.
Этот принцип имеет большое практическое и лечебное значение, а также представляет большой теоретический интерес. Он означает, например, что если мигрень необходима для поддержания физиологического и эмоционального баланса больного, то приступы будут продолжаться, несмотря на исключение из системы тех или иных ее частных звеньев. Можно удалить одну височную артерию или орган-мишень, но их функцию возьмут на себя другие артерии и органы. Можно заблокировать действие серотонина, но приступы возобновятся с использованием иных промежуточных механизмов.
Такие функциональные системы (говорит Лурия), сложные по своему строению, пластичные в разнообразии своих элементов, обладающие свойством динамической ауторегуляции, очевидно, являются правилом управления деятельностью человека.
Резюме
На нижнем функциональном уровне мигренозная реакция характеризуется длительным усилением парасимпатического или трофотропного тонуса, причем ей предшествует противоположное состояние, которое, кроме того, следует за приступом. На высшем функциональном уровне приступ мигрени характеризуется активацией (и последующим угнетением) бесчисленных корковых полей – от первичных сенсорных областей до наиболее сложных интегративных областей. Мигрень считается медленно развивающейся формой центрального мозгового пароксизма. В случае ауры длительность приступа в 20—200 раз длиннее эпилептического припадка; приступ же простой мигрени превосходит длительность эпилептического припадка в тысячи раз. Необходимо также считать мигрень сложной адаптивной задачей, решаемой сложной функциональной системой, в которой средства исполнения (отличающиеся чрезвычайно большой вариабельностью) подчинены выполнению главной задачи.
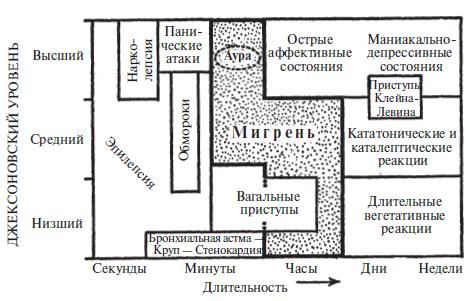
Рис. 9. Мигрень и некоторые сопутствующие расстройства. Мигрень и сопутствующие ей расстройства представлены здесь в координатах временной оси и оси нейронных уровней (джексоновских уровней). Характеристики обеих модальностей подвержены изменениям – отсюда метаморфозы, к которым так склонна мигрень. Все представленные здесь расстройства имеют отчетливые признаки, тем не менее у каждого из них есть пограничные зоны, которые могут сливаться с такими же зонами других расстройств.
Мигрень переходит в диффузную пограничную область сопутствующих пароксизмальных и адаптивных реакций. Схематически это можно представить с помощью двух координатных осей: по ординате отложены функциональные (джексоновские) уровни, а по абсциссе – длительность реакций (рисунок 9). В идеале должна присутствовать и третья ось, на которой следует отметить различные синдромы мигрени и сопутствующие реакции в соответствующее время и на соответствующем функциональном уровне.
Для практических целей (диагностических и лечебных) мы можем идентифицировать большую часть случаев мигрени как мигрень и ничего больше. Теоретически мы можем ожидать развития разнообразных гибридных и переходных приступов на тех уровнях, где мигрень сливается с эпилепсией, обмороками, вагальными приступами и другими вегетативными и аффективными кризами [46]. Время от времени мы сталкиваемся с такими гибридными и переходными приступами, которые делают нас беспомощными в плане дифференциальной диагностики (превращая ее в полностью бессмысленное занятие).
Можно думать, что физиологические границы очерчены отнюдь не более отчетливо, чем границы клинические, и, может быть, поэтому химерической мечтой является стремление отыскать в нервной системе уникальный и патогномоничный «мигренозный процесс». Мы не можем представить себе, что какое-то очередное исследование позволит нам выявить какую-то особую патологию, такую же кристально прозрачную и четкую, как сочетание волн и спайков на ЭЭГ больного эпилепсией. Вместо этого надо ожидать широкий спектр нервной активности, проявляющейся в разные временные моменты на разных функциональных уровнях.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
§29. Ассоциации и их физиологическая основа
§29. Ассоциации и их физиологическая основа Запоминая какие-либо образы, мысли, слова, чувства, движения, мы всегда запоминаем их в определённой связи друг с другом. Без установления тех или других связей невозможно ни запоминание, ни узнавание, ни воспроизведение.Что
§52. Физиологическая основа чувств
§52. Физиологическая основа чувств У человека, как мы знаем, в основе всех психических процессов лежат нервные процессы, протекающие в коре больших полушарий. Чем же с физиологической стороны отличается чувство от других психических процессов?Всякий психический процесс
СТАДИЯ ПЕРВИЧНЫХ ВЗАИМОДЕЙСТВИЙ: ФИЗИОЛОГИЧЕСКАЯ ПРЕЛЮДИЯ К ОБЪЕКТНЫМ ОТНОШЕНИЯМ
СТАДИЯ ПЕРВИЧНЫХ ВЗАИМОДЕЙСТВИЙ: ФИЗИОЛОГИЧЕСКАЯ ПРЕЛЮДИЯ К ОБЪЕКТНЫМ ОТНОШЕНИЯМ Большинство исследователей согласны с тем, что мы рождаемся подготовленными для участия во взаимодействии. Сандлер (1975) концептуально описывает это взаимодействие как часть биологически
Физиологическая природа эмоций
Физиологическая природа эмоций Вот вам и эволюция человеческого мозга. Женщина XXI века все реже плачет от несчастной любви, предпочитая мимолетные связи. Мужчину же травмирует просмотр фильма или отказ во взаимности. Что это? Новые реалии мышления или закономерное
Измена налицо, и он утверждает, что имела место чисто физиологическая неверность
Измена налицо, и он утверждает, что имела место чисто физиологическая неверность Нужно убедиться, что всей этой истории пришел конец. Это обязательно. От этого зависит дальнейшая тактика поведения.Если ты точно знаешь, что измена была лишь однажды, то можешь принять его
Часть I Проявления мигрени
Часть I Проявления мигрени Введение Первая проблема возникает с самим словом «мигрень», так как этот термин предполагает существование головной боли (в половине головы) как основной характеристики заболевания. Необходимо с самого начала сказать, что головная боль
2 Эквиваленты мигрени
2 Эквиваленты мигрени Обзор множества симптомов, составляющих в совокупности клиническую картину простой мигрени, показывает, что заболевание нельзя отождествлять с каким-то одним из этих симптомов. Проявления мигрени складываются из бесчисленного множества
5 Структура мигрени
5 Структура мигрени Мы рассмотрели главные типы картин мигрени во всем их невообразимом разнообразии и неоднородности. Теперь нам следует остановиться, чтобы как следует разобраться в этих картинах и по возможности их упростить. По мере того как мы углубляемся в предмет,
Часть II Эпидемиология мигрени
Часть II Эпидемиология мигрени Введение Многие больные считают, что мигрень возникает спонтанно и без причины. В научном плане такой взгляд ведет к абсурду, в эмоциональном – к фатализму, в терапевтическом – к бессилию. Мы должны принять, что у всех приступов мигрени
6 Предрасположенность к мигрени
6 Предрасположенность к мигрени Мы привыкли думать, что любой частный ответ является либо заученным, либо врожденным, что является причиной ошибок в суждениях о таких вещах… Является ли ответ унаследованным или приобретенным? Ответ: ни тем, ни другим. Да или Нет –
Часть III Основа мигрени
Часть III Основа мигрени Введение Мы рассмотрели формы, в которых может проявляться мигрень, и условия, при которых она может возникнуть. Большая часть этих сведений была известна Лайвингу, Говерсу и Хьюлингсу Джексону сто лет назад, и эти врачи – и прежде всего Лайвинг –
10 Физиологические механизмы мигрени
10 Физиологические механизмы мигрени Те, кто не понимает истории, обречены на ее повторение. Сантаяна Историческое введениеПятую часть своего шедевра Лайвинг посвятил рассмотрению и критическому разбору множества теорий мигрени, существовавших и в его время, а затем
13 Психологические подходы к изучению мигрени
13 Психологические подходы к изучению мигрени Это большая ошибка нашего времени… что врачи отделяют душу от тела. Платон В предыдущей главе мы коснулись некоторых примитивных реакций, которые могут пролить свет на происхождение и дифференцировку мигренозных реакций у
Часть IV Способы лечения мигрени
Часть IV Способы лечения мигрени Введение Теперь мы могли бы сказать, что, прояснив природу мигрени – во всяком случае, насколько мы смогли это сделать, – мы закончили свою миссию и можем с чувством выполненного долга поставить точку. Можно даже сказать, что говорить о
14 Общие лечебные мероприятия при мигрени
14 Общие лечебные мероприятия при мигрени Мы знаем, что приблизительно одна десятая часть населения страдает простой мигренью, одна пятидесятая часть – классической мигренью, и очень малое число больных страдает редкими вариантами мигрени (мигренозная невралгия,
16 Последние достижения в лечении мигрени
16 Последние достижения в лечении мигрени Тот факт, что приступы мигрени могут провоцироваться внезапными эмоциями или возникать в ответ на напряженный образ жизни, или быть следствием личностных особенностей индивида, часто дает повод считать мигрень «функциональным»