Память
Память
Кора головного мозга содержит более 10 млрд нейронов. Нейроны коры имеют в среднем от 10 000 до 100 000 связей. Количество и сложность связей увеличиваются с возрастом. Изначально, при рождении, большая часть нейронов «нового мозга» не осуществляет какой-либо функции. Такая «девственная» структура служит единственной цели — быть готовой к запоминанию всего, что станет происходить с человеком в течение жизни. Каждый нейрон в течение жизни «зафиксирует» на себе некую картину, которая будет соответствовать текущей на тот момент картине активности мозга. Впоследствии такой нейрон будет в состоянии узнать повторение «зафиксированной» картины и в случае своей активации — воспроизвести картину, соответствовавшую моменту запоминания.
В 2000 году американский нейробиолог Эрик Кандел получил Нобелевскую премию по медицине «За открытие молекулярных механизмов работы синапсов». Память изучалась им на примере формирования условного рефлекса у гигантского моллюска — морского зайца аплизии (Aplysia).
Моллюску осторожно трогали сифон, и тотчас вслед за этим сильно били по хвосту. После такой процедуры моллюск некоторое время реагировал даже на легкое прикосновение к сифону бурной защитной реакцией, но вскоре все забывал (кратковременная память). Если «обучение» повторялось несколько раз, формировался стойкий условный рефлекс (долговременная память).
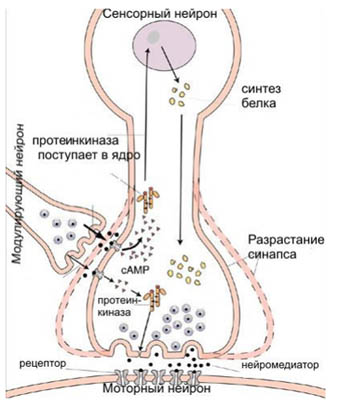
Рисунок 18. Молекулярный механизм работы синапсов.
На рис.18 показаны два синапса. Первый служит для передачи импульса от сенсорного нейрона к моторному. Второй синапс передает импульс от модулирующего нейрона к окончанию сенсорного. Если в момент прикосновения к сифону модулирующий нейрон «молчит» (по хвосту не бьют), в синапсе 1 выбрасывается мало нейромедиатора и моторный нейрон не возбуждается.
Однако удар по хвосту приводит к выбросу нейромедиатора в синапсе 2, что вызывает важные изменения в поведении синапса 1. В окончании сенсорного нейрона вырабатывается сигнальное вещество cAMP (циклический аденозинмонофосфат). Это вещество активизирует регуляторный белок протеинкиназу А. Протеинкиназа А, в свою очередь, активизирует другие белки, и это в конечном счете приводит к тому, что синапс 1 при возбуждении сенсорного нейрона (то есть в ответ на прикосновение к сифону) начинает выбрасывать больше нейромедиатора, и моторный нейрон возбуждается. Это и есть кратковременная память: пока в окончании сенсорного нейрона много активной протеинкиназы А, передача сигнала от сифона к мышцам жабры и чернильного мешка осуществляется более эффективно.
Если прикосновение к сифону сопровождается ударом по хвосту много раз подряд, протеинкиназы А становится так много, что она проникает в ядро сенсорного нейрона. Это приводит к активизации другого регуляторного белка — транскрипционного фактора CREB. Белок CREB «включает» целый ряд генов, работа которых в конечном счете приводит к разрастанию синапса 1 (как показано на рисунке) или к тому, что у окончания сенсорного нейрона вырастают дополнительные отростки, которые образуют новые синаптические контакты с моторным нейроном. В обоих случаях эффект один: теперь даже слабого возбуждения сенсорного нейрона оказывается достаточно, чтобы возбудить моторный нейрон. Это и есть долговременная память.
Остается добавить, что, как показали дальнейшие исследования, у высших животных и у людей память основана на тех же принципах, что и у аплизии (Han, и др., 2007).
Эрик Кандел расшифровал устройство памяти моллюска. Он показал, что достаточно трех нейронов, чтобы реализовать базовую модель его поведения.
Конечно, память человека устроена значительно сложнее. С моей точки зрения, самую красивую теорию организации сложной памяти выдвинул А. Н. Радченко. Если говорить максимально упрощенно, то своим открытием он показал организацию ассоциативной памяти на рецепторных кластерах (Радченко, 2002). Чтобы понять, что это значит, вспомним сначала о ритмах мозга.
В 20-х годах немецкий психиатр Ганс Бергер, изучая электрическую активность головного мозга человека, впервые обнаружил слабые колебания с частотой около 10 Гц и назвал их альфа-волнами. Их амплитуда составляет всего около 30 миллионных долей вольта. Так был открыт альфа-ритм — наиболее четкий образец электрической активности мозга. Изучение этих еле заметных волн выросло в новый раздел науки, называемый электроэнцефалографией — ЭЭГ (Berger, 1929).
В возбужденном состоянии нейроны обладают импульсной активностью. Причем эта активность не индивидуальна для каждого нейрона, а синхронизирована с активностью других. Наблюдаемые ритмы мозга — это проявление суммарной активности нейронов. Каждый всплеск на энцефалограмме — это след совместного «залпа» нейронов. Активность нейронов имеет групповой характер, в каждой группе задается свой ритм импульсов. То, что мы наблюдаем при простом подсоединении электродов к голове,— это сигнал сложной формы, образованный суммированием ритмов различной частоты.
Из сигнала сложной формы выделяют отдельные гармонические составляющие. Достаточно четко прослеживаются несколько основных ритмов:
- дельта-ритм (от 0,5 до 4 колебаний в секунду, амплитуда — 50—500 мкВ);
- тета-ритм (от 5 до 7 колебаний в секунду, амплитуда — 10—30 мкВ);
- альфа-ритм (от 8 до 13 колебаний в секунду, амплитуда — до 100 мкВ);
- сигма-ритм — «веретена» (от 13 до 14 колебаний в секунду);
- бета-ритм (от 15 до 35 колебаний в секунду, амплитуда — 5—30 мкВ);
- гамма-ритм (от 35 до 100 колебаний в секунду, амплитуда — до 15 мкВ).
Английский нейрофизиолог и пионер робототехники Грей Уолтер и
американский психолог Уоррен Мак-Каллок высказали достаточно обоснованную гипотезу о том, что альфа-ритм характеризует процесс внутреннего «сканирования» мысленных образов при сосредоточении внимания на какой- нибудь умственной проблеме. Наблюдается, например, совпадение между частотой альфа-волн и периодом инерции зрительного восприятия (примерно 0,1 секунды) (Walter, 1967).
Зачем нужна такая синхронная импульсная активность? Чтобы понять это, надо посмотреть, как устроена поверхность нейрона. Поверхность нейрона вся покрыта клеточными рецепторами. Рецепторы — это белковые молекулы, соединяющие нейрон с внешней средой и служащие проводниками информации.
Клеточный рецептор — участок клеточной оболочки, молекулярная структура которого характеризуется избирательностью к некоторым определенным веществам (напр., антигенам, вирусным рецепторам), способностью вступать с ними в химическое взаимодействие (присоединять, связывать) (Кузина, и др., 2006).
Рецепторы можно объединять по схожести реакций в группы — кластеры. Количество кластеров на поверхности нейрона — 104 — 105.
Импульсная активность других нейронов передается через синапсы на поверхность каждого нейрона и образует «интерференционную картину», причем каждому состоянию соответствует определенная картина «пучностей».
Интерференция волн — сложение в пространстве двух (или нескольких) волн, при котором в разных точках получается усиление или ослабление амплитуды результирующей волны. И характерна для всяких волн независимо от их природы: для волн на поверхности жидкости, упругих (например звуковых) волн, электромагнитных (например радио- или световых) волн (Горелик, 1959).
Пучность — участок стоячей волны, в котором колебания имеют наибольшую амплитуду (Горелик, 1959).
За счет синхронности излучения нейронов картина пучностей повторяется каждый раз, когда повторяется картина их активности, то есть пучности появляются каждый раз на тех же самых местах на поверхности нейрона. Рецепторные кластеры могут фиксировать такие пучности и являться впоследствии элементами, которые обеспечивают активацию нейрона в случае повторения ситуации. И, наоборот, группа нейронов, будучи активирована, может привести к «восстановлению» окружающей картины.
Синхронная активность делает возможным образование на поверхности нейрона устойчивой интерференционной картины. Устойчивой в том смысле, что при повторении ситуации активности нейронов за счет совпадения фаз их импульсов интерференционная картина на поверхности нейрона повторится. Из такого понимания роли ритмов мозга несложно развить рассуждения относительно их роли в процессах запоминания и распознавания.
Описанная модель памяти относится к разряду «голографических», так как по своему принципу напоминает устройство голограммы, где запоминание объемного изображения достигается за счет фиксации интерференционной картины, возникающей при использовании когерентных источников освещения.
Вспомним устройство коры головного мозга.
Первичная проекционная зона сосредотачивает в себе всю информацию, поступающую извне. Нейроны, располагающиеся непосредственно над проекционной зоной и имеющие связи в одном из ее участков, способны фиксировать «воспоминания», относящиеся к этому участку. Это могут быть воспоминания о каких-либо звуках, зрительных образах, совершенных движениях и т. п. Поднимаясь выше, мы получаем нейроны, которые фиксируют более сложные картины, состоящие из более комплексных воспоминаний. Важно отметить, что при обратной активации, то есть воспоминании, сигналы доходят до первичной проекционной коры и вызывают вполне конкретные образы.
Исходя из такой пространственной организации уместно говорить о зонах мозга с точки зрения относительной локализации воспоминаний, относящихся к какой-либо специальной области деятельности: о зрительной зоне, слуховой зоне и так далее.
Вспомним об опытах Пенфилда по раздражению различных участков коры мозга. Посмотрим на это в свете модели текущего представления и понятий внутреннего языка.
При раздражении участков, где находятся проекционные зоны зрительной коры, пациент наблюдает элементарные образы: линии, точки, вспышки, движение точки слева направо... Это и есть базовые понятия внутреннего языка.
При воздействии на вторичную ассоциативную зону зрительные образы усложняются: возникают образы людей, людей в той или иной одежде, людей в статике или движении. Это соответствует более сложным понятиям.
Воздействие электрических импульсов на третичные зоны приводит к активации воспоминаний о конкретных событиях, которые строятся, в свою очередь, на использовании понятий вторичной ассоциативной коры.
Очевидно, что, поднимаясь по коре все выше, мы имеем дело со все более комплексными воспоминаниями, построенными на воспоминаниях предыдущего уровня.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Память
Память В когнитивной психологии под памятью понимают психический процесс приобретения, хранения, удержания и воспроизведения информации. Механизм памяти представляет собой три взаимосвязанных процесса: кодирование, хранение и воспроизведение
4.4. Память
4.4. Память Понятие памяти. Все то, что человек когда-то воспринимал, не исчезает бесследно – в коре больших полушарий головного мозга сохраняются следы от процесса возбуждения, которые создают возможность повторного возникновения возбуждения в отсутствие вызвавшего его
Память? А что память?
Память? А что память? «Существует одна тревога, связанная со старением, без которой, я думаю, мы можем прекрасно обойтись: когда мы не можем вспомнить имя или что мы собирались сделать… Это не значит, что мы превращаемся в сумасшедших». * * *Вы можете обнаружить, что ваша
Рабочая память и кратковременная память
Рабочая память и кратковременная память Многие считают, что понятие «рабочая память», которое сейчас так активно используется, запустил в научный обиход психолог Алан Бэддели в начале 1970-х годов[30]. Он предложил разделить рабочую память на три блока. Один отвечает за
Часть I. Как в два раза улучшить память за сорок пять минут, или Введение в голографическую память
Часть I. Как в два раза улучшить память за сорок пять минут, или Введение в голографическую память С чего все начиналось… Несколько лет назад, после окончания последнего занятия по развитию памяти, один из студентов предъявляет претензии в отношении результатов
Память
Память Прежде чем продолжить свой анализ того, что Субъекты, находящиеся в состоянии гипноза, видят в Мире Душ, я хотел бы предоставить больше информации о категориях памяти и ДНК. Есть люди, которые Убеждены, что все воспоминания хранятся в ДНК. Таким образом они
Память прошлого и память будущего
Память прошлого и память будущего Мои коллеги-психологи, исследователи памяти, предполагают, что резервы нашей памяти практически неисчерпаемы. Нашей головы хватит, чтобы мы запоминали все и всегда: и вон тот случайный разговор на улице, и колыхание каждой ветки вон того
Память
Память Как отмечалось ранее, IQ является одной из популярнейших зависимых переменных психологических исследований старения. Память — еще одна популярная переменная. В 1991-1993 годах 34 % статей, опубликованных в журналах Psychology and Agingn Journal of Gerontology: Psychological Science, так или иначе
«Повседневная» память и долговременная память
«Повседневная» память и долговременная память Рассмотрим еще два вопроса, имеющих отношение к теме «Память». До сих пор основное внимание уделялось стандартным лабораторным методам, часто исполь-зуемым при изучении памяти в любой возрастной период. Два последних
§ 6. Память
§ 6. Память Память — интегрированное психическое отражение прошлого взаимодействия человека с действительностью, информационный фонд его жизнедеятельности.Способность хранить информацию и избирательно ее актуализировать, использовать для регулирования поведения —
Память
Память Прежде чем произведение будет создано, писатель должен подготовить для него необходимый материал. Он долго наблюдает за окружающей действительностью, многое переживает в собственной жизни. Впечатления внешнего и внутреннего мира образуют в совокупности
5 Память
5 Память ВОПРОСЫ О ПАМЯТИ Тест 1. Запоминание во время заучивания Ниже приводится список слов. Быстро прочитайте все слова по порядку по одному разу, затем откройте с. 68 и впишите все слова, которые сумели запомнить. Не пропустите ни одного слова при чтении. Для уверенности
Память
Память Темно в долинах памяти, словно в осеннем сумеречном лесу, где в печальной наготе застыли стволы деревьев, а яркие цветы и буйная зелень листьев обратились в шуршащий под ногами ковер. Нет пути назад. Никакие заклятия не вдохнут жизнь в отцветшие сады, в отзвучавшие